Protein Birincil Yapısı
Kısaca: Peptit ve proteinlerin birincil yapısı, bu moleküllerin yapı birimleri olan amino asitlerin doğrusal sırası, veya daha genel olarak, bir proteini oluşturan atomlar arasındaki kovalent bağların spesifikasyonudur. ...devamı ☟
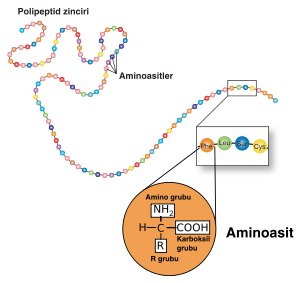
Protein yapısı
3 yıl öncegrubu bir peptit bağı içinde yer almaz. Canlılarda bir proteinin birincil yapısı o proteini kodlayan gen tarafından belirlenir. DNA'daki belli bir nükleotit...
Protein ikincil yapısı
3 yıl öncebazen fark edilebilir. Katlanma (kimya) Protein birincil yapısı Protein üçüncül yapısı Protein dördüncül yapısı Translasyon Yapısal motif ^ Richards F...
Protein
3 yıl öncediğer moleküllerden yüzlerce kere daha büyüktü. Bu yüzden proteinlerin kimyasal yapısı (birincil yapıları) 1949'a kadar önemli bir araştırma konusuydu, o...
Protein, 1838, 1926, 1958, 1962, ATP, Akciğer, Alanin, Albümin, Alem, Amfibi, Biyoloji, Hücre, CanlıBiyomoleküler yapı
3 yıl önceBiyomoleküler yapı biyomoleküllerin (başlıca protein, DNA ve RNA'nın) yapısıdır. Bu moleküllerin yapısı genelde birincil, ikincil, üçüncül ve dördüncül yapı olarak...
İnsülin
3 yıl öncedizilimi (birincil yapı) ise İngiliz moleküler biyolog Frederick Sanger tarafından belirlenmiştir ve yapısı tamamen çözümlenebilen ilk protein insülin olmuş...
Vazopressin
7 yıl önceüzere memelilerin büyük çoğunluğunda bulunan bir hormondur. Vasopressin'in birincil görevi, böbreklerden su geri emilimini arttırmaktır. Vasopressin, peptid...
Taşıyıcı RNA
7 yıl önceamino asidi taşıyabilir. tRNA'nın birincil, ikincil ve üçüncü yapıları vardır. İkincil yapısı genelde yonca yaprağı yapısı olarak gösterilir. Üçüncül yapı...
Hücre iskeleti
3 yıl öncebüyüme veya küçülme yeteneğine sahip üç ana proteinden oluşan dinamik bir yapıdır. Hücre iskeletinin yapısı, işlevi ve dinamik davranışı organizma ve hücre...